C O A C H I N G
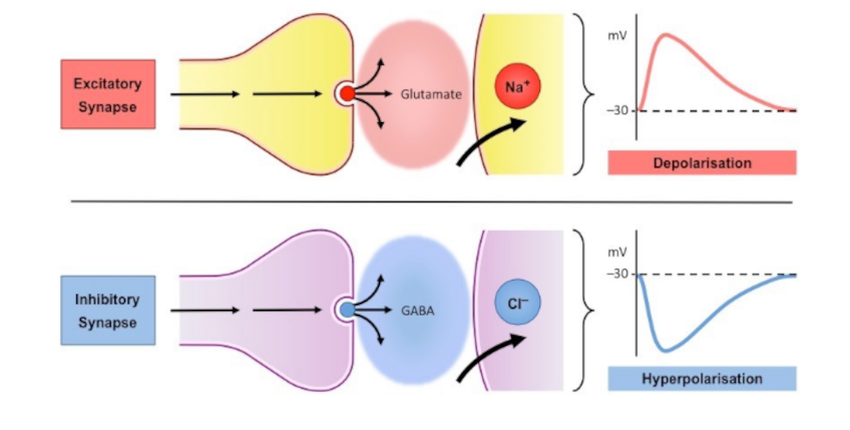
Glutamate
Introduction
Neurons are what we call the cells inside the brain that produce and respond to neurotransmitters, and other chemical messengers. Most neurons have a storage of neurotransmitters contained inside of them, inside of vesicles. When a neuron is at “rest”, meaning it is not releasing its stored neurotransmitters, it is referred to as being inhibited. In order to excite a neuron it must surpass a voltage threshold, which allows for different ions, specifically calcium, to enter into the cell and allow vesicles containing neurotransmitters to fuse with the cellular membrane and release the stored neurotransmitters. The neurotransmitters are released out of the neuron into the synaptic space where they can bind to a receptor located on another neuron. The synaptic space is simply the space between two communicating neurons. When the voltage surpasses a specific threshold, the vesicle is capable of fusing into the cellular membrane and releasing the neurotransmitters stored inside. To be slightly less abstract, the resting potential of a neuron is estimated to be around -70 millivolts. This means, that inside the neuron is 70 millivolts lower than the environment outside of the neuron. These numbers are quantified according to the charge of all the molecules within a neuron. It is estimated that when a neuron surpasses the voltage of 50 millivolts, it becomes capable of releasing the stored neurotransmitters.
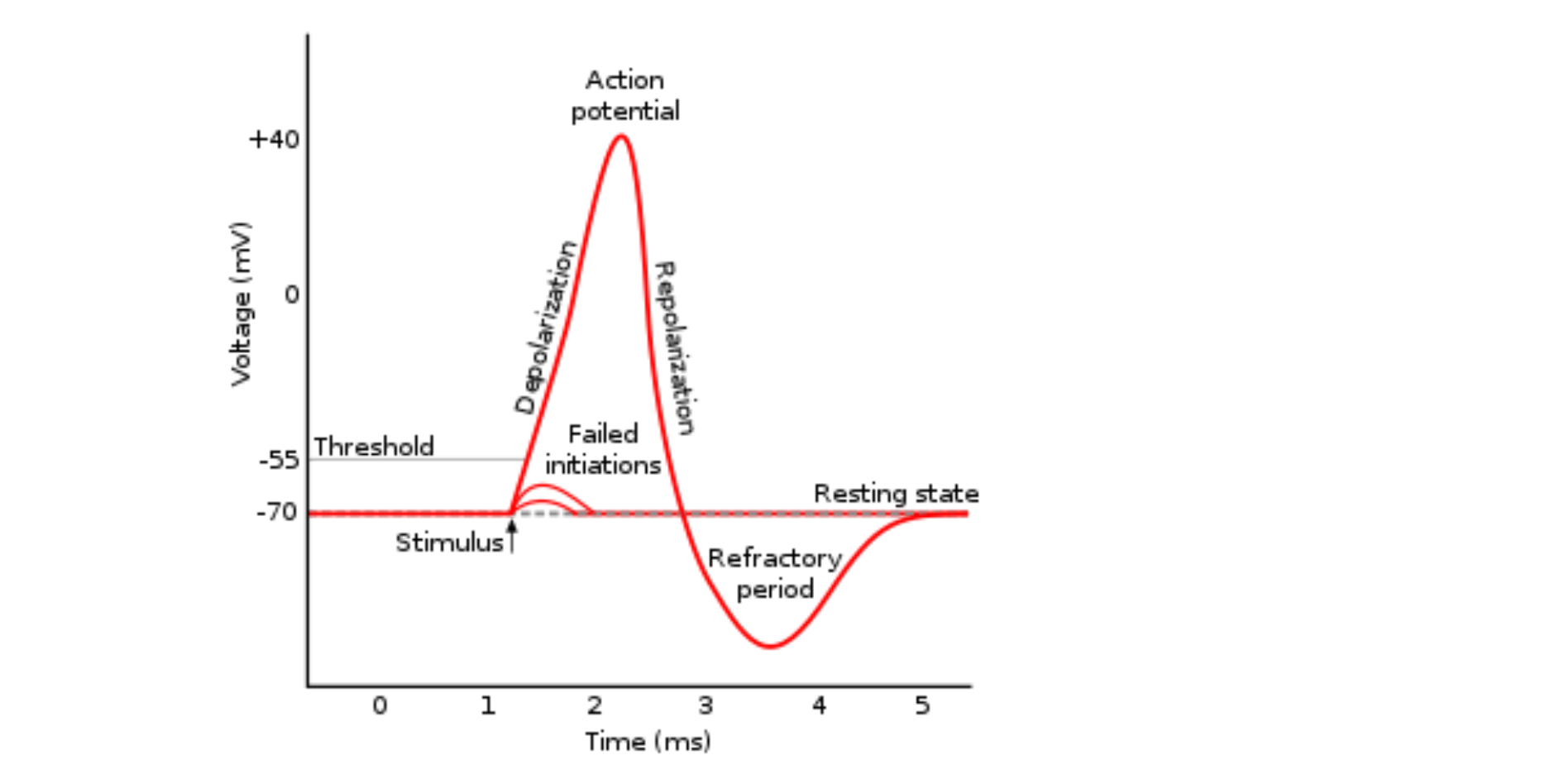
Within the nervous system, we have broad classes of neurons referred to as excitatory and inhibitory neurotransmitters. Excitatory neurotransmitters are neurotransmitters that assist the neuron in reaching its voltage threshold by increasing the amount of positively charged molecules that enter the neuron. Conversely, inhibitory neurotransmitters typically cause positively charged ions to leave the neuron, and negatively charged ions to enter, thus decreasing the voltage of the neuron and making it more difficult to reach its voltage threshold.
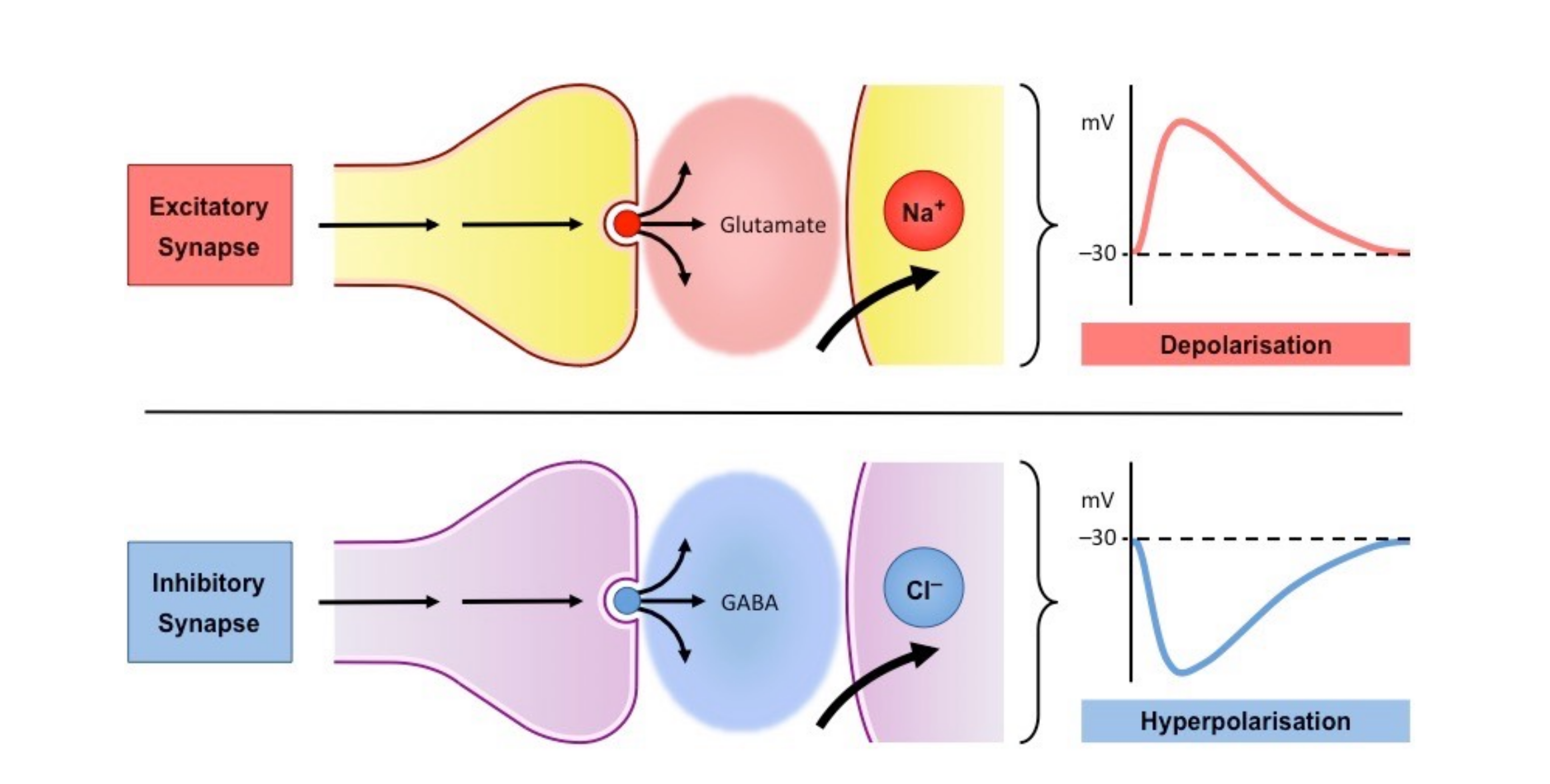
If you haven't yet, check out my article all about the primary inhibitory neurotransmitter GABA. In today's discussion, we will talk all about the primary excitatory neurotransmitter glutamate.
Production and storage
Glutamate is secreted by neurons located in the central nervous system. The neurons in the spinal cord that store and release glutamate are referred to as the presynaptic glutamatergic neurons. These neurons store and release hourmate into the area known as the synapse. Remember that the synapse is simply the space between two communicating neurons. The neuron that is inhibited by glutamate is referred to as the postsynaptic neuron.
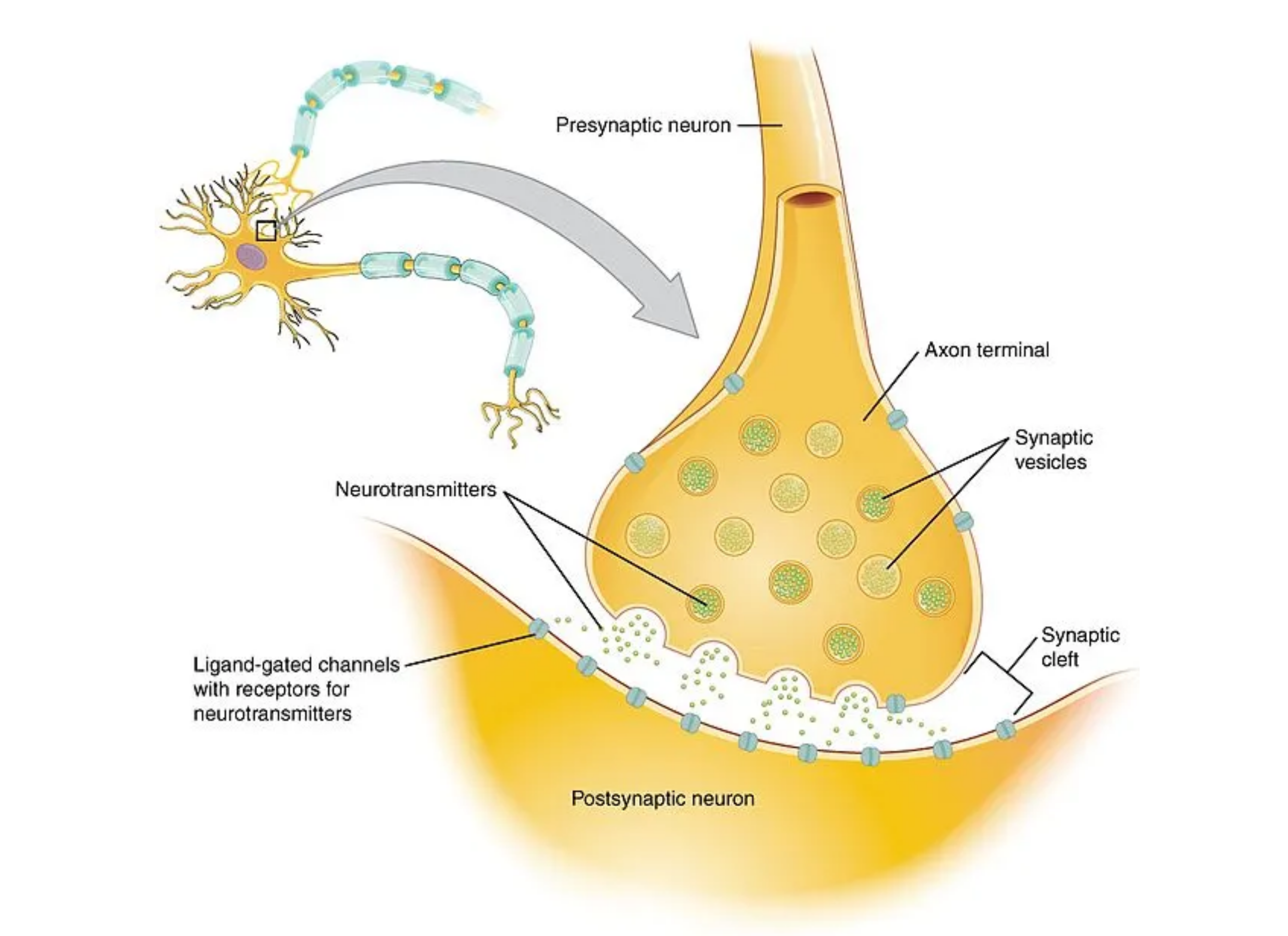
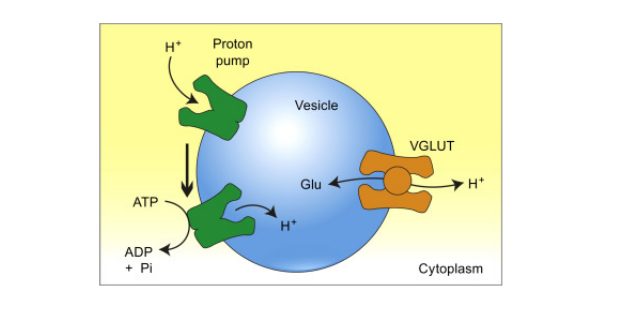
Before we get into the effects of glutamate, let's talk about how we get glutamate into the secretory vesicle within the presynaptic neuron in the first place. Interestingly, glutamate isn't actually able to be uptaken into the neuron in order to be stored. Thus, neurons need to uptake a different molecule out of the bloodstream called glutamine, which is eventually converted to glutamate. Glutamine can enter through a group of amino acid transporters called sodium coupled neutral amino acid transporters (SNATs). Glutamine is then converted into glutamate via the glutaminase enzyme (1).
Glutamatergic neurons contain VGLUT vesicles within them. These vesicles were named based upon the protein transporter present on their membrane called vesicular glutamate transporters (VGLUT) on their membrane. In addition, they contain another transporter called vacuolar hydrogen ATPase. This protein utilizes ATP, removing hydrogen from ATP. This hydrogen is used as a substrate for the antiporter on the membrane referred to as VGLUT. An anti-porter is simply a protein channel that will update one molecule while subsequently releasing a stored molecule. This anti-porter will release the stored hydrogen back out of the vesicle and uptake the free glutamate.
In order to use glutamate, it must be released from the V-GAT vesicle, into the synapse. This is stimulated through a series of mechanisms that increase the concentration of positively charged molecules inside of the glutamatergic neurons, thus causing the neuron to surpass the voltage required to release the stored glutamate. This is referred to as excitation, and it said exactly the opposite of what glutamate does! This process begins when sodium channels located along the axon of the neuron open. The sodium molecules are positively charged, and they increase the voltage of the neuron. This will stimulate voltage gated calcium channels to change shape. As the name implies, the activity of these channels is dictated by the voltage of the neuron. When the neuron reaches a specific voltage these calcium channels change shape and allow calcium influx into the neuron.
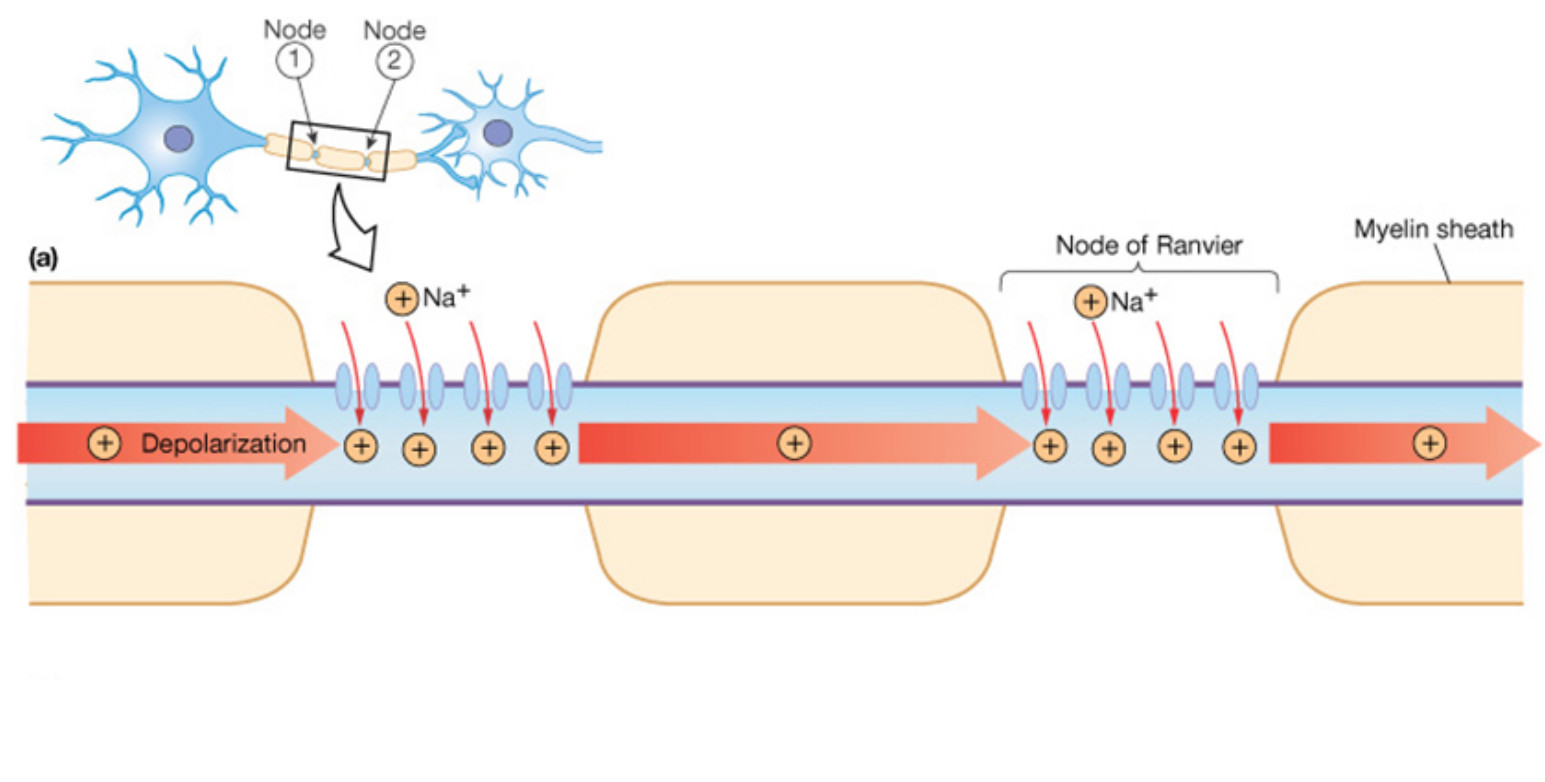
Calcium is the substrate that allows the vesicle containing glutamate to fuse the plasma membrane and release the glutamate stored inside of the vesicle. This is mediated by proteins present on the vesicle and on the plasma membrane referred to as SNARE proteins. Calcium allows these proteins to attach to one another, essentially fusing the vesicle into the membrane, and release the stored neurotransmitter, which in this case is glutamate.
Calcium is necessary for allowing the vesicle to fuse to the membrane of the neuron, and release glutamate outside of the neuron. Calcium allows for the interaction of proteins called SNARE proteins, located on the membrane of the vesicle, and the membrane of the neuron to bind to one another and fuse the vesicle to the membrane.
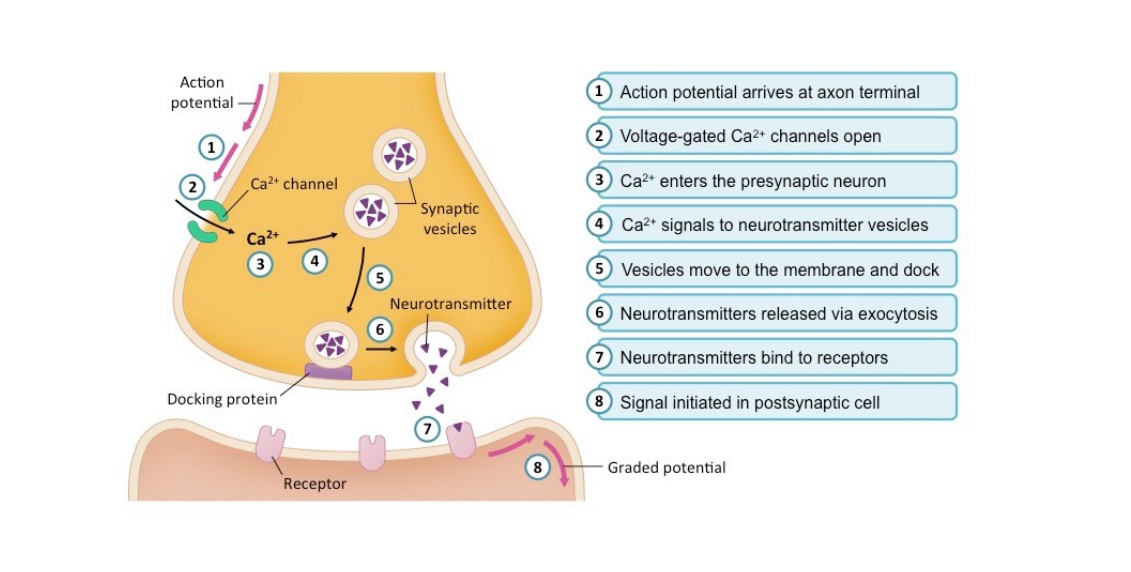
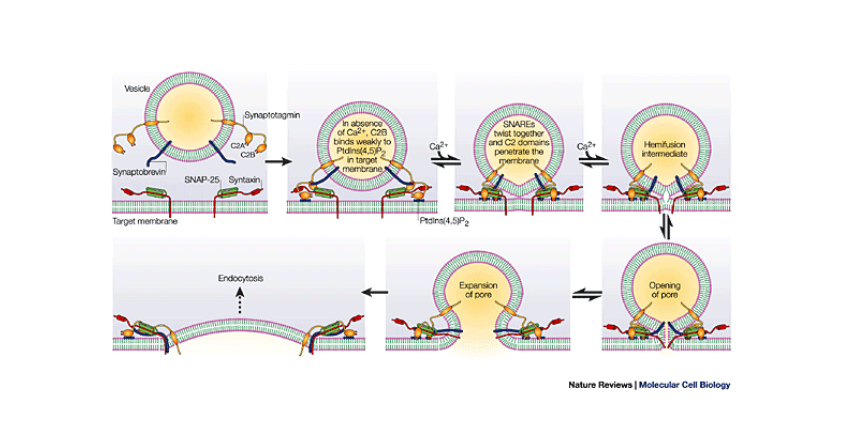
If you're a nerd like me, here is the cellular physiology of this process:
On the membrane of the vesicle, there is a protein called synaptobrevin. Also, on the membrane of the vesicle is another protein called synaptotagmin. On the membrane of the neuron, where the vesicle will dock, there’s a protein called syntaxin. Additionally, on the membrane, there is a protein called SNAP-25. Simply, these three proteins form a complex that allows the vesicle to dock on the membrane, until it is ready to be fused into the membrane and undergo exocytosis. Exocytosis is simply the process of releasing distorted glutamate from the vesicle into the external environment. Here is where calcium comes in. On the protein we mentioned earlier called synaptotagmin, there are two distinct portions. The C2A portion and the C2B portion. Calcium binds to the C2A portion on the synaptotagmin, then the C2B region will see use into the membrane around the SNARE proteins. While it is slightly unclear the exact mechanism, the synaptotagmin protein causes the membrane of the neuron to begin to curve inwards in the direction of the vesicle. As the membrane of the neuron and the vesicle are both made out of the same phospholipid material, the membranes simply join together, and release the glutmate into the external environment.
Glutamate receptors
Once glutamate is released, it binds to a receptor on the postsynaptic neuron. There are two types of glutamate receptors, ionotropic receptors and metabotropic receptors.
Ionotropic
There are 3 distinct ionotropic receptors, NMDA, AMPA, and kainate receptors. All three of these receptors work to allow positively charged molecules into the neuron, which stimulates the same process as I walked through above to allow storage vesicles to bind to the membrane and release their stored neurotransmitters.
Attached to the receptors are positive ion channels. These receptors often coexist with one another on the membrane of postsynaptic neurons. That is, most port synaptic glutamate neurons contain multiple different types of glutamate receptors. The kainate and AMPA receptors are the weakest, and most easily activated. These receptors allow for sodium to enter into the cell, which as described above lead to the downstream activation of sodium gated calcium channels, and neurotransmitter release.
The NMDA channel is Very interesting. The NMDA receptor is much stronger than the former two receptors, however it is also much more difficult to activate. The reason is because this receptor often has a molecule of magnesium between the proteins that should allow positively charged ions to enter into the cell when glutamate binds. With that being said, as sodium begins to enter into the cell due to stimulation of the NMDA and kainate receptors, this magnesium can actually be dislodged, allowing for a high abundance of positively charged ions to enter into the neuron, thus allowing it to reach its voltage threshold. This is why I mentioned earlier that oftentimes there are many glutamate receptors on the same neuron, because glutamate has the ability to regulate the function of its own receptors. These channels all work in tandem to raise the voltage of the cell, and the result is the neuron surpassing 50 millivolt, allowing for voltage gated calcium channels to open and the neurotransmitters that are contained within the neuron, although through the same process to be released as glutamate did.
Metabotropic receptor
Metabotropic receptors for glutamate are G protein coupled receptors. Simply, the outside of the portion of the receptor on the outside of the cell contains the binding site, and on the inside it is attached to what is called a g stimulatory protein. Attached to the G stimulatory protein is a molecule of GDP which inhibits the protein. Upon binding, the GDP is released, and GTP binds to the G stimulatory protein. GTP simply has one extra phosphate than GDP, which is used to phosphorylate other molecules. The process of phosphorylation is simply when a phosphate group is added to some sort of intracellular protein. This phosphate is subsequently used as energy to either inhibit or activate a given intracellular protein. In this case, GTP phosphorylates an enzyme called phospholipase C (PLC).The PLC enzyme converts a protein in the cell called PIP2 into two new proteins called DAG and IP3. DAG is used for important cellular functions, but we want to focus our attention on IP3. IP3 binds to what is called an IP3 calcium channel in an organelle inside of the cell called the endoplasmic reticulum where calcium is stored. The result is the opening of the calcium channel, and influx of calcium out of storage into the rest of the cell. Thus, the calcium can allow for vesicle fusion and release (2).
Nutritional considerations and closing remarks
As mentioned, the NMDA receptor is very strong, and is inhibited by magnesium. Insufficiency and magnesium could result in excessive excitation of neuronal connections, which can cause symptoms associated with anxiety, ADHD, and increase the likelihood of sleep disturbances. Thus, adequate magnesium in the diet can be an extremely potent tool for possibly helping to mitigate the likelihood of the onset of these abnormalities (3).
Magnesium supplements can come in a myriad of different forms of magnesium. Some magnesium supplements, such as magnesium oxide, or poorly absorbed therefore will have very minimal effect on raising the body's magnesium status. Magnesium glycinate is extremely well absorbed, however it's important to note that much dietary and supplemental magnesium will not be able to get into the brain because it cannot be transported through the blood brain barrier (4). However, it does appear that magnesium threonate has the most potency for increasing brain levels of magnesium. However, increasing magnesium intake in general, from any source, is almost always a beneficial idea unless you already have uncommonly high magnesium levels (5).
Knowing the glutamate is an excitatory neurotransmitter, and can also be found in our food sometimes scares people. Thus, they try to limit glutamate consumption whenever possible. Importantly, this is not necessary. The reason is because just like magnesium, glutamate cannot just freely pass through the blood brain barrier into the brain. Which is why I mention that the glutamate that is stored inside vesicles is originally created from glutamine. Thus, limiting the consumption of things such as monosodium glutamate, or glutamate rich foods isn't necessary for someone who may have Symptoms of excess glutamate.
Another important caveat is that we do not want to always try and inhibit the production of glutamate. Glutamate is vital for proper learning, and memory. When you go to make neurons become activated, they actually up regulate the number of glutamate receptors located on the membrane, which is referred to as postsynaptic potentiation. This potentiation makes a presynaptic and postsynaptic neuron much more sensitive to glutamate stimulation, therefore much more likely to become activated. So the influence of glutamate, and many other neurotransmitters, is potentiation is actually how we are able to learn new things, and remember important information (6).
With these caveats out of the way, I hope you found all of this information interesting, however do not take this as a comprehensive overview of everything to do with glutamate. We are learning more and more about the brain, influence of neurotransmitters, and the function of neurotransmitters on a daily basis. Thus, if you discover anything that I may have left out, I absolutely encourage you to reach out to me so I can update this article as often as possible!
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4919214/ https://www.ncbi.nlm.nih.gov/books/NBK10802/#:~:text=Several%20types%20of%20ionotropic%20glutamate,receptors%20(Figure%207.11C). https://pubmed.ncbi.nlm.nih.gov/30807974/#:~:text=The%20evidence%20from%20this%20meta,to%20further%20investigate%20this%20issue. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6683096/ https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9786204/ https://www.frontiersin.org/articles/10.3389/fphar.2012.00182/full

Recent Blog
 01
01
Show at the University
 02
02
Show at the University
 03
03
Show at the University
 04
04